|
Una
visión nítida del mundo visual que nos rodea requiere de la producción
de movimientos oculares.
 Los
movimientos oculares constituyen una estrategia motora que contribuye
de tres modos distintos a evitar la pérdida de información visual: Los
movimientos oculares constituyen una estrategia motora que contribuye
de tres modos distintos a evitar la pérdida de información visual:
-
- Estabilizando el mundo visual en movimiento mediante
el mantenimiento de la mirada sobre puntos fijos en el espacio. Esto
es posible debido a la existencia de dos movimientos oculares reflejos,
el vestíbulo-ocular y el optocinético.
- -
Orientando la mirada y explorando el medio externo, llevando
y manteniendo la mirada hacia determinados blancos visuales, gracias
al sistema de movimientos rápidos, denominados sacádicos y al sistema
de vergencia.
-
Fijando sobre la zona de mayor percepción visual la imagen de
objetos que se desplazan en el espacio por medio del sistema de seguimiento.
Aunque
la producción de todos los tipos de movimientos oculares requiere de
un conjunto enorme de estructuras nerviosas, la organización a nivel
troncoencefálico recoge las rutas finales para la mayor parte de los
movimientos oculares.
Durante
los últimos 15 años nos hemos centrado en la caracterización de los
grupos neuronales que, implicados en la generación de los movimientos
oculares en el plano horizontal, se encuentran localizados en el mesencéfalo,
el puente y el bulbo.
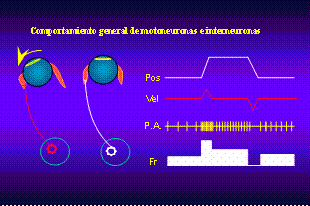
 Así
se han caracterizado, en el animal despierto y alerta, la actividad
de neuronas identificadas como motoneuronas e interneuronas internucleares
en los núcleos motores del III y VI par craneal. Estas neuronas constituyen
la salida motora del sistema nervioso central responsable de la producción
de los movimientos oculares en el plano horizontal. El comportamiento
de las mismas se caracteriza por la producción de una frecuencia de
potenciales de acción que es proporcional a la posición y velocidad
del ojo en la órbita. Así
se han caracterizado, en el animal despierto y alerta, la actividad
de neuronas identificadas como motoneuronas e interneuronas internucleares
en los núcleos motores del III y VI par craneal. Estas neuronas constituyen
la salida motora del sistema nervioso central responsable de la producción
de los movimientos oculares en el plano horizontal. El comportamiento
de las mismas se caracteriza por la producción de una frecuencia de
potenciales de acción que es proporcional a la posición y velocidad
del ojo en la órbita.
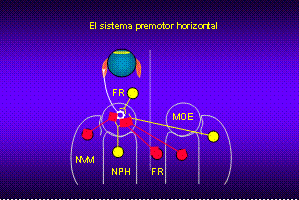
También se ha caracterizado
la actividad de los grupos neuronales de la protuberancia y el bulbo
que hacen sinapsis con las motoneuronas e interneuronas, es decir las
neuronas premotoras. Para la identificación de las neuronas premotoras
se utrilizó, en el animal despierto, la técnica del promediado de potenciales
de campo extracelulares inducido por una neurona aislada. Esta técnica
permite identificar si existe o no conexión sináptica con un núcleo
determinado, el carácter excitador o inhibidor de la sinapsis y la actividad
de la neurona durante movimientos oculares espontáneos o inducidos por
distintas estimulaciones sensoriales. Concretamente se han identificado
y registrado las neuronas de los núcleos vestibulares mediales (NVM),
las neuronas de la formación reticular (FR) pontina y bulbar y las neuronas
del núcleo prepositus hipoglossi (NPH). Cada grupo de neuronas mantiene
una conexión recíproca excitadora e inhibidora con el núcleo motor.
 De manera muy esquemática, se puede decir que las neuronas de
la formación reticular codifican la velocidad ocular durante los movimientos
oculares rápidos, las neuronas vestibulares codifican la velocidad de
la cabeza y las neuronas del NPH la posición ocular durante cualquier
tipo de movimiento ocular. El descubrimiento de las propiedades de descarga
de las neuronas del NPH en relación a la posición ocular, el hecho de
que este núcleo recibe una copia de cada uno de los grupos neuronales
que codifican velocidad y la existencia dentro de este núcleo con neuronas
con distintos perfiles de sensibilidad a la velocidad y a la posición
ocular nos hizo proponer que en este núcleo podría tener lugar la transformación
de las señal de velocidad en señal de posición. Apuntando en esta dirección,
se obtuvo que la regularidad de las neuronas del NPH era muy superior
a la de cualquier otra neurona premotora con señal de posición ocular
y comparable a la de las motoneuronas y que la probabilidad de descarga
de una motoneurona, con respecto a la de una neurona con señal de posición,
incrementa a tiempos monosinápticos, mientras que el aumento de probabilidad
para una neurona con señal de velocidad-posición tiene lugar a tiempos
di- o trisinápticos. Este conjunto de resultados nos hizo proponer un
modelo en cascada para la generación de la señal de posición en el NPH.
De manera muy esquemática, se puede decir que las neuronas de
la formación reticular codifican la velocidad ocular durante los movimientos
oculares rápidos, las neuronas vestibulares codifican la velocidad de
la cabeza y las neuronas del NPH la posición ocular durante cualquier
tipo de movimiento ocular. El descubrimiento de las propiedades de descarga
de las neuronas del NPH en relación a la posición ocular, el hecho de
que este núcleo recibe una copia de cada uno de los grupos neuronales
que codifican velocidad y la existencia dentro de este núcleo con neuronas
con distintos perfiles de sensibilidad a la velocidad y a la posición
ocular nos hizo proponer que en este núcleo podría tener lugar la transformación
de las señal de velocidad en señal de posición. Apuntando en esta dirección,
se obtuvo que la regularidad de las neuronas del NPH era muy superior
a la de cualquier otra neurona premotora con señal de posición ocular
y comparable a la de las motoneuronas y que la probabilidad de descarga
de una motoneurona, con respecto a la de una neurona con señal de posición,
incrementa a tiempos monosinápticos, mientras que el aumento de probabilidad
para una neurona con señal de velocidad-posición tiene lugar a tiempos
di- o trisinápticos. Este conjunto de resultados nos hizo proponer un
modelo en cascada para la generación de la señal de posición en el NPH.
 En
la actualidad seguimos trabajando en este modelo y dado que las neuronas
de los núcleos vestibulares también presentan cierta señal de posición
ocular estamos interesados en conocer de donde proviene dicha señal
y que valor funcional tiene. En este sentido estamos trabajando en la
modificabilidad que pueden sufrir las señales de posición en ambos núcleos
y su posible papel en la compensación vestibular tras lesión. En
la actualidad seguimos trabajando en este modelo y dado que las neuronas
de los núcleos vestibulares también presentan cierta señal de posición
ocular estamos interesados en conocer de donde proviene dicha señal
y que valor funcional tiene. En este sentido estamos trabajando en la
modificabilidad que pueden sufrir las señales de posición en ambos núcleos
y su posible papel en la compensación vestibular tras lesión.
También
hemos caracterizado algunos de los neurotransmisores que intervienen
en las sinápsis aferentes al núcleo prepositus y en el
procesamiento interno en dicho núcleo. Entre ellos se encuentran
los neurotransmisores clásicos, como el glutamato que actua tanto
sebre receptores NMDA como no NMDA, la glicina, el GABA, la acetilcolina
y el óxido nítrico. el óxido nítrico no
parece formar parte de manera directa de la generación de la
señal de posición, pero por el contrario parece controlar
las entradas de velocidad desde los núcleos vestibulares. El
control de dichas entradas es muy importante, porque como ambos núcleos
vestibulares mantienen una descarga tónica sobre el núcleo
prepositus en condiciones de inmovilidad, la integración para
generar una señal de posición proporcional a la velocidad
de desplazamiento requiere una eliminación de dicha entradas
tónicas. Así, hemos propuesto que el óxido nítrico
controla la actividad de la propia neurona que lo produce mediante el
control de la liberación de GABA desde la neurona vestibular.
Este papel fisiológico es importante además porque ayuda
a controlar las diferencias de entradas que se producen sobre el núcleo
prepositus hipoglossi desde los núcleos vestibulares, por ejemplo,
como consecuencia de una lesión vestibular o cervical, ayudando
a disminuir el vertigo que se produce como consecuencia.
La
acetilcolina, por el contrario, parece intervenir directamente, a través
de los recptores muscarínicos, en la generación de la
señal de posición ocular . La microinyección local
de antagonistas colinérgicos provoca una incapacidad para mantener
la posición del ojo en la órbita, alterando la capacidad
del núcleo para generar la señal de posición. En
la actualidad estamos caracterizando las neuronas colinérgicas
del núcleo prepositus y su posible interacción con las
neuronas nitrérgicas.
|
-
Escudero, M. y Delgado-García, J.M. "Behavior of reticular, vestibular
and prepositus neurons terminating in the abducens nucleus of the
alert cat", Exp. Brain Res., 71: 218‑222, 1988.
-
De la Cruz, R.R., Escudero,M. y Delgado- García, J.M. "Behavior
of medial rectus motoneurons in the alert cat". European
J. Neuroscience,1:
288‑295, 1989.
-
Gómez, C., Quero, J.M. y Escudero, M. "Computer simulation of
the neural discharge carried by the abducens nerve during eye fixation
in the cat". Int. J. Biomedical Computing, 24:207-215,
1989.
-
Delgado-García, J.M.,Evinger, C., Escudero, M.y Baker R., "Behavior
of accesory abducens motoneurons in the alert cat", J.
Neurophysiol., 64:413‑422, 1990.
-
Escudero, M., de la Cruz, R.R. y Delgado-García, J.M. "A physiological
study of vestibular and prepositus hypoglossi neurones projecting
to the abducens nucleus in the alert cat", J.Physiol.(Lond.),458:539-560,1992.
-
Cheron, G., Mettens, P. and Escudero, M. "Effects of APV
injection into the abducens and the prepositus hypoglossy on
the generation of eye position signal" en "Information
Processing Underling Gaze Control", J.M. Delgado-García,
Godaux, E. and P.P. Vidal (eds.), págs.: 21-31, Elsevier Science
Ltd, Oxford.1994.
-
Cheron, G., Escudero, M. y Godaux, E. "Behavior of the brainstem
neurons projecting to the flocculus in the alert cat. I. Medial vestibular
nucleus". J. Neurophysiol., 76:17591774, 1996.
-
Escudero, M., Cheron, G., y Godaux, E. "Behavior of the brainstem
neurons projecting to the flocculus in the alert cat.
I. Prepositus hypoglossi nucleus". J. Neurophysiol., 76:1775-1785,
1996.
- Moreno-López,
B. Escudero, M., Delgado-García, J.M., Estrada, C.
Nitric oxide production by brain stem neurons is required for normal
performance of eye movements in alert animals.
Neuron,
17: 739-745, 1996.
-
Moreno-López, B., Estrada, C. y Escudero, M. Mechanisms
of action and targets of nitric oxide in the oculomotor system.
J. Neurosci.,
18(24):10672-10679, 1998.
-
Moreno-López, B., Escudero, M., de Vente, J. y Estrada, C.
Morphological
identification of nitric oxide sources and targets in the cat oculomotor
system.
J. Comp. Neurol. 435: 311-324, 2001.
- Moreno-López,
B., Escudero M. y Estrada C.,
Nitric oxide facilitates gabaergic neurotransmission in the oculomotor
system: a physiological mechanisms in eye movement control.
J. Physiol., 540.1: 295-306, 2002.
- Navarro-López,
J.D., Alvarado, J.C., Márquez-Ruiz, J., Escudero, M., Delgado-García,
J.M. y Yajeya, J., A cholinergic synaptically triggered event participates
in the generation of persistent activity necessary for eye fixation,
J. Neurosci. 24(22):5109-5118, 2004.
|

